Disturbed regeneration of saplings of Korean fir (Abies koreana Wilson), an endemic tree species, in Hallasan National Park, a UNESCO Biosphere Reserve, Jeju Island, Korea
Department of Forestry, Environment, and Systems, Kookmin University, Seoul 02707, Republic of Korea kimeuns@kookmin.ac.kr
Department of Biological and Environmental Science, Dongguk University, 32, Dongguk-ro, Ilsandong-gu, Goyang-si, Gyeonggi-do 10326, Republic of Korea
Graduate School, Dongguk University, 32, Dongguk-ro, Ilsandong-gu, Goyang-si, Gyeonggi-do 10326, Republic of Korea
Korea National Park Research Institute, Korea National Park Service, Wonju-si, Gangwon-do 26441, Republic of Korea
Graduate School, Kookmin University, Seoul 02707, Republic of Korea
Graduate School, Kookmin University, Seoul 02707, Republic of Korea
Institute of Forest Science, Kookmin University, Seoul 02707, Republic of Korea
Institute of Forest Science, Kookmin University, Seoul 02707, Republic of Korea
Graduate School, Kookmin University, Seoul 02707, Republic of Korea
Graduate School, Kookmin University, Seoul 02707, Republic of Korea
Department of Forest Products and Biotechnology, Kookmin University, Seoul 02707, Republic of Korea
Abstract
Limited knowledge is available on the regeneration of Korean fir (Abies koreana Wilson), an endemic plant species, growing on the upper part of Mt. Hallasan, a volcanic mountain, located in the central part of Jeju Island, Korea. A forest stand with the size of 1 ha dominated by Korean fir trees was established and all the trees with DBH 2 cm or larger were mapped and surveyed. Initial analysis indicated that the numbers of saplings with their DBHs between 2 cm and 10 cm were very small and that there was a big gap in the frequency of the number of saplings regenerated from the forest stand. It seems clear that the regeneration of the Korean fir trees was disturbed for longer than the last two decades, potentially by the browsing of the seedlings by ungulate including Siberian roe deer and by the physical hindrance of the dwarf bamboo to the development of the saplings of the Korean fir. Urgent measures and extensive studies are needed to promote the natural regeneration of the tree species on the dynamics of the forest regeneration and the mechanism of forest development of the forests on the Mt. Hallasan, Jeju Island, Korea.
Keywords
Decline, Dwarf bamboo (Sasa quelpaertensis Nakai), Frequency distribution, JEJU Island, Korean fir (Abies koreana Wilson), Mt. Hallasan, Regeneration, Saplings, Seedlings, Siberian roe deer (Capreolus pygargus Pallas), Stand dynamics, Ungulate browsing
Introduction
Jeju Island is the southern-most and the largest island in Korea. It is a volcanic island and is designated from UNESCO as a UNESCO World Biosphere Reserve in 2002, World Natural Heritage in 2007 and UNESCO Global Geopark in 2010 (Woo et al., 2013). Mt. Hallasan, located in the central part of the island, has the highest peak in Korea with its altitude of 1950 m above the mean sea level.
On the upper part of the mountain, Korean fir (Abies koreana Wilson), a subalpine coniferous tree species, is growing forming forest stands. Korean fir is an endemic tree species classified by Wilson (1920) and categorized as an endangered species by IUCN (http://www.iucnredlist.org/details/31244/0). In addition, Korean fir was designated as Climate-sensitive Biological Indicator Species (CBIS) in Korea (Lee et al., 2010; Park et al., 2015). The forests of Korean fir trees suffered from a severe decline in 1960s, whose causes were not known until now (Kim, 1994; 1996). Recently, the Korean fir forests on Mt. Hallasan suffer from decline, which has diverse symptoms including: the big Korean fir trees fallen due to higher susceptibility for windfall, especially situated on a lava floor, where soil is not deep enough to fully support the root systems of the tall trees in height; patches of dead trees of Korean fir of different sizes, whose causes of the death not well studied. To cope with the societal needs to address the problems of the decline of the Korean fir tree species, in late October 2015, the Warm Temperate and Subtropical Forest Research Center of the Korea Forest Research Institute organized an International Symposium to conserve the Korean fir trees on Mt. Hallasan in Seogwipo City, Jeju Island, Korea. Presentations on managerial issues to prevent the decline of the tree species as well as botanical characteristics of the species were made and discussion was extended to the measures to prevent the decline of the tree species on Mt. Hallasan (Korea Forest Research Institute, 2015).
In Jeju Island, there is a saying from the local people that Korean fir can live only one hundred years, which is a quite interesting saying possibly based upon long-term observation of nature in Jeju Island. This saying gives an insight to the decline of the big and old trees of Korean fir, which indicates that it is an unavoidable natural phenomenon related to increased vulnerability of the trees with shallow root systems standing on thin soil layers formed on lava floor of basalt. Vulnerability of the aged trees to the ever powerful winds, consequently lowered vigors of the trees and increased susceptibility to harsh and changing environment disturbed by omnipotent typhoons and frequent droughts might be the causes for the decline of the old trees. In other part of the world, the phenomena of declining fir trees at high altitudes were reported as the series of waves of dying fir trees, so called, ‘fir waves’ or ‘Shimagare’ (Sprugel, 1976; Kohyama, 1988).
A worrisome phenomenon that we can observe from the forests on Mt. Hallasan is the fact that it is quite difficult to observe small trees of Korean fir trees at forest stands on Mt. Hallasan, which is a real concern for us, ecologists, to observe at the forest indicating the eventual decline of the tree species in the long run. From stand dynamics’ view points of the natural forests, trees are regenerated from seeds or sprouts from stumps and roots, which develop into seedling, saplings, and eventually into maturing trees (Meyer, 1952; Helms, 1998; Halpern, 2001). Saplings are differentiated from seedlings in size, especially in height. Although there are different measures to differentiate sapling from seedling, saplings are generally regarded as the small trees whose heights are taller than certain heights (e.g., 1.3–1.5 m, 2.0 m, or breast height), whereas seedlings are smaller trees whose heights are smaller than certain heights. As the saplings are the small trees that can be developed into the bigger and mature trees at the forests, they are the indicators of the future condition of the forest, in composition, health, development, etc. If there are few saplings in the forests where bigger trees are simultaneously under decline, we can evaluate that the forests are under the condition of real decline.
In conserving the Korean fir forests, managing the forests sustainably, and, ultimately, preventing the decline of the Korean fir forests on Mt. Hallasan, it is crucial to establish and promote the natural regeneration of saplings of the Korean fir trees on the forests. Currently, there are two major factors that potentially disturb the regeneration of the forests, which are the browsing of the seed germinates, young seedlings, and growing saplings by the Siberian roe deer (Capreolus pygargus Pallas); and the physical hindrances and competition of the dwarf bamboo (Sasa quelpaertensis Nakai) inhibiting the establishment of the seedlings of Korean fir on Mt. Hallasan. There are many cases supporting the effects of the factors around the world. In October 2015, IUFRO International Conference was held to discuss the effects of ungulate browsing on forest regeneration and silviculture, which was organized by IUFRO and Swiss Federal Institute for Forest. Halpern (2001) reviewed the literature on the constraints on the conifer regeneration. In order to establish the successful seedlings and saplings, there are series of stages where the constraints to be overcome from seed production, seed dispersal, seed germination, seedling establishment, sapling establishment, and eventually mature trees. On affecting the regeneration, he suggested that there are groups of factors including abiotic and biotic variables, where biotic variables include herbivore browsing, competition with other plants, mycorrhizal limitation, fungal pathogens and phytotoxic effects.
There are few studies carried out to investigate the factors related to the regeneration of the saplings of Korean fir on Mt. Hallasan, Jeju Island. Currently, we rarely know about what has been happening at the forests on the animal-plant interactions, interspecific competing among plant species, adaptation mechanism of plant species surviving the harsh environmental conditions of the forest stands. On this issue of the decline of the Korean fir forests, it is important to address such questions as:
- –
- what are the general patterns of frequency distribution over tree sizes (DBH Class) of Korean fir trees?
- –
- how much has the pattern of frequency distribution disturbed and what does it imply in interpreting the situation of the decline of Korean fir forest?
- –
- what would it imply in the management of the decline of Korean fir forests?
Here, emphasis is placed upon finding out the gaps in frequency distribution of the Korean fir trees on Mt. Hallasan, Jeju Island, Korea. The objectives of this study are (1) to investigate the current status of frequency distribution of the saplings of the Korean fir trees, (2) to characterize the gap between the expected frequency distribution and the actual frequency distribution of the number of Korean fir trees, (3) to extend ecological interpretation of the gap to the potential causes for the disturbed regeneration, and (4) to suggest the measures to prevent disturbance and to promote the regeneration of the Korean fir trees on Mt. Hallasan, Jeju Island, Korea.
Materials and methods
Study area and sites
For the purpose of Long-Term Ecological Research (LTER), an area of 5 sites with each site with the size of 1 ha was established near the area of Jindallaebat Shelter at a sub-alpine forest on Mt. Hallasan, Jeju Island, Korea (Fig. 1). The area is located on the eastern slope of Mt. Hallasan at a location with altitude between 1520 m and 1640 m above the mean sea level.
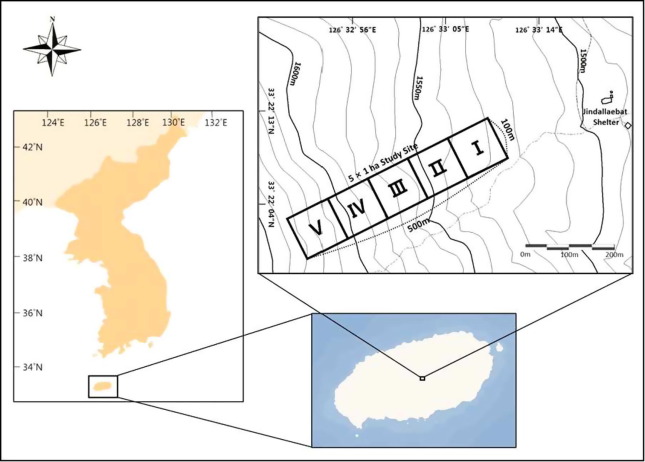
Site survey and data
At the study area, the data surveyed from Site I with the size of 1 ha (100 m × 100 m) were used for the analysis and discussion for this study.
Although there are different ways to define seedlings and saplings, seedling is defined as “a small tree germinated from seed” and a sapling is defined as “a usually young tree larger than a seedling but smaller than a pole, whose size varies by region” (Helms, 1998; Kerr et al., 2002; Kerr and Mackintosh, 2012). For example, in forest management in US, saplings are regarded as trees between the DBH (Diameter at Breast Height) Classes of 1″–4″, which implies that the saplings are the trees with their heights at least taller than breast height (about 1.3–1.4 m above the ground) with their DBH. For data analysis, we applied 2 cm size class in DBH and regarded the trees as saplings with their DBHs between 2 cm and 10 cm. In this study, the trees included in the 3 cm DBH Class are the those with their DBHs larger than or equal to 2 cm and those with their DBHs smaller than 4 cm. Similarly, the trees included in 5 cm DBH Class are trees with their DBHs larger than or equal to 4 cm and those with their DBHs smaller than 6 cm. In the surveying of the trees for this study, the trees with their DBHs smaller than 2 cm were not tallied.
Using the data obtained from the survey, information was analyzed on the frequency distribution of the trees by DBH class, stand statistics of the sites including density of trees and basal area were calculated for the forest stand.
Results and discussion
Stand statistics and frequency distribution of tree species by DBH Class
At the study site (Site I), for the mature and dominant trees with their DBHs equal to or larger than 20 cm, the number of the trees for the Korean fir tree species (265 trees, 92.3%) is much higher than those for Japanese yew (Taxus cuspidata Siebold et Zucc.; 2 trees, 0.7%) and deciduous hardwood species (20 trees, 7.0%), which indicates that Korean fir is the dominant trees species in the forests (Table 1). For the trees with their DBHs in between 11 cm and 19 cm, the number of the trees for the Korean fir tree species (288 trees, 57.3%) is much higher than that for Japanese yew (28 trees, 5.6%) and also higher than that of deciduous hardwood species (187 trees, 37.2%), which indicates that deciduous hardwood species are trying to outcompete under the dominant Korean fir trees.
| DBH class in cm | Species | |||
|---|---|---|---|---|
| Korean fir | Japanese yew | Deciduous hardwood species | Total | |
| 2–10 | 69 (19.5%) | 97 (27.5%) | 187 (53.0%) | 353 (100%) |
| 11–19 | 288 (57.3%) | 28 (5.6%) | 187 (37.2%) | 503 (100%) |
| ⩾20 | 265 (92.3%) | 2 (0.7%) | 20 (7.0%) | 287 (100%) |
| Total | 622 (54.4%) | 127 (11.1%) | 394 (34.5%) | 1143 (100%) |
The cases for the saplings or small trees growing under the canopy are quite contrasting compared with those for the dominant and co-dominant trees. For the saplings trees with their DBHs in between 2 cm and 10 cm, the number of the Korean fir trees (69 trees, 19.5%) and that for Japanese yew (97 trees, 27.5%) are similar each other but much smaller than that compared to the deciduous hardwood species (187 trees, 53.0%), which indicates that deciduous hardwood species are occupying more densely the lower layer of the forest stand than the trees of Korean fir and Japanese yew. It is striking to see that there are no sapling of Korean fir with their DBHs smaller than 3 cm and very few saplings of Korean fir trees with the DBH of 4 cm and 5 cm. Still, for the saplings with their DBHs equal to or smaller than 5 cm, the number of trees for Korean fir is the lowest compared with those of Japanese yew and the other deciduous tree species. This indicates that succession is taking place at this forest stand of Korean fir to be replaced by the stands of deciduous tree species of Korean cherry (Prunus maximowczii Rupr.), Silverly mountain ash (Sorbus commixta Hedl.), Erman’s betula (Betula ermanii Cham.), Korean whitebeam (Sorbus alnifolia (Siebold et Zucc.) K. Koch) and Mongolian oak (Quercus mongolica Fischer), as well as by the stand of Japanese yew in the long-run.
Gaps in the regeneration of saplings and small understory trees at the site
Considering the variability of the density of the trees growing at different sites with different soil conditions, aspects, topography, stand history, etc., it is safe to assume that the interpretation could be limited in making direct comparisons of statistics drawn from different study sites carried out at different forest stands on Mt. Hallasan. As there were a few results showing the figures of the stand densities of the Korean fir trees, we would like to make comparisons of our results with the observations from other published studies. Kim and Choo (2000) reported that Korean fir trees with their sizes smaller than 10 cm in DBH Class were about 37% (155 trees among 419 Korean fir trees) from the data they sampled in 1996 at a different forest stand on Mt. Hallasan. Kim et al. (2007a,b) reported that Korean fir trees with their sizes smaller than 10 cm in DBH Class were about 40% (242 trees among 606 Korean fir trees) from the data they sampled at another forest stand on Mt. Hallasan. Compared with the tree densities observed from the sites of the studies above, the portion of the density of the small trees from this study Site (24.4%) is much smaller than those of the other studies stated above. In contrast, Koh et al. (1996) reported that, although there were variations in the number of seedlings among the sites, the trees with their heights in 2–3 m and their diameters at root collar in 8–10 cm were most dominant in the forest stand, which may indicate that there were relatively much more small trees than those observed from the forest stand of this study carried out in recent years. The results of these reports indicated that the smaller trees were not many about 20 years ago, but not so small in numbers observed from this study.
At this study site, the number of the saplings for the tree species is quite small and even smaller for the small saplings with their DBH Classes of 3 cm, 5 cm, and 7 cm. It should be noted that there is a big gap in the number of saplings of the tree species, especially, for the Korean fir trees between the expected number of trees and actual number of the trees at the study site. If the population of Korean fir trees develops under normal conditions without major disturbances, there will be much higher number of saplings than that of the current ones. It is quite difficult to suggest any number as the definite estimates for the expected numbers of saplings for the DBH Classes because it can vary depending on the site conditions and biotic and abiotic factors that affect them at the sites, however.
Fig. 2 is the diagram for frequency distribution for the number of trees for Korean fir, Japanese yew and other deciduous hardwood species by DBH class at the study site near Jindallaebat Shelter area on Mt. Hallasan, Jeju Island, Korea.
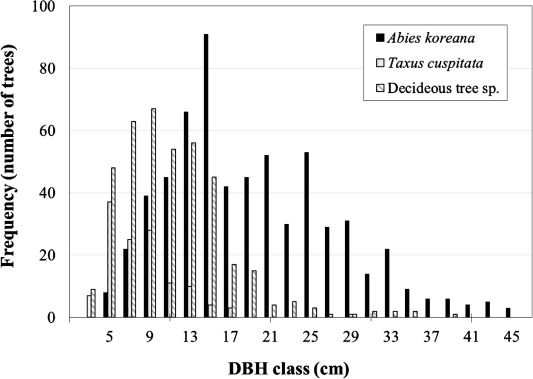
From ecological view points of the natural forest, trees are regenerated from seeds or sprouts from stumps and roots, which develop into seedlings and, further, saplings, and eventually into maturing trees. As the saplings are the small trees that can be developed into the bigger trees at the forests, they are the indicators of the future condition of the forest, in composition, health, development, etc. If there are few saplings in the forests where bigger trees are simultaneously under decline, we can evaluate that the forests are under real decline.
Generally, in uneven aged natural forests, the number of trees decreases exponentially by the increase of tree size such as diameter and height. As a result, the frequency distribution of trees shows reverse-J shaped distribution by the increase of tree sizes, especially with the DBH increase (Meyer, 1952; Lorimer, 1980; Leak, 1996; Loewenstein et al., 2000; Wang et al., 2000; Ducey, 2006; Rubin et al., 2006; Westphal et al., 2006). In order to describe the gap more objectively, the cumulative frequency was drawn against DBH Class for the site and an arbitrary reverse-J distribution curve was superimposed on the figure, when the gap in the frequency distribution was more clearly shown (Fig. 3). In Fig. 3, it is apparently shown that the numbers of the saplings of Korean fir, Japanese yew, and other deciduous hardwood species are much less than those of expected frequency that can be seen from the natural forest stands without being affected by the major disturbance factors.
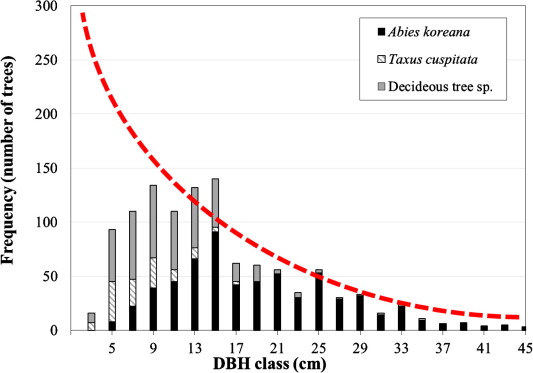
This gap of low frequency for the number of the saplings of trees, especially for the Korean fir trees, strongly suggests that there are some factors that disturbed the natural processes of regeneration of seedlings and saplings. This is one of the most important findings of this study, which provide significant implication that this gap would lead to the ultimate decline of the forests if no measures are to be taken to promote the regeneration of seedlings and saplings of the Korean fir tree species from the forests on Mt. Hallasan, Jeju Island, Korea. Here, the success for establishing enough number of the seedlings and saplings of the Korean fir tree species can be used as one of the most important indicators to prevent the decline of the tree species from the forest stands.
Another important finding to note is that the saplings of the Korean fir trees are under severe competition with the saplings of other tree species including Japanese yew and deciduous hardwood species. Therefore, it is quite possible to find that the dominance of Korean fir trees can be replaced by dominance of such deciduous hardwood species when the tall trees of Korean fir die and decline due to some disturbance factors such as typhoons, droughts, etc. at the site. This means that if current conditions prevail and no significant measures are taken to promote natural regeneration of the Korean fir trees, this forest stand of Korean fir will be replaced by the forest stand of hardwood tree species.
Sapling gaps for more than last 20 years at the Korean fir Forest Stands
With these phenomena of sapling gaps considered, it can be described that the forest stands have been disturbed in the past for the period from when seeds germinate to when the saplings develop to mature trees at the forests. It is difficult to suggest finite number in years for the time for the seeds to grow beyond saplings because it can differ depending on the fertility and micrometeorology of the site, genetic characteristics of individual seeds, climate regimes that affect the development of the seeds into the seedlings, saplings and beyond the stages of saplings including such environmental factors as droughts, strong winds (typhoons), damping-off pathogens, etc. Considering the swallow depth and low fertility of the soils at the forests, we can postulate that it takes more than twenty years for the seeds to develop into new saplings with the heights above the breast (about 1.3–1.4 m high). Considering all these, we maintain that the regeneration of the Korean fir saplings has been disturbed for more than 20 years in the past at the forest stands.
Among many factors that can affect the regeneration of the Korean fir trees on Mt. Hallasan, Jeju Island, Korea, there are two distinct candidate factors that can disturb the regeneration of the trees at the forests, which are browsing of seedlings by the Siberian roe deer and the severe competition of dwarf bamboo interrupting the regeneration of the tree species from the initial stage of seed germination.
Two important time periods for the successful regeneration of saplings of Korean fir
To provide ideas on the time period for the seeds to develop into seedlings and saplings, and eventually to mature trees on Mt. Hallasan, we made a schematic diagram showing the stand dynamics of the Korean fir trees from the seed germination on the ground at the site to the maturing stages of the Korean fir trees at the forest stand with a relative scale in tree height on y axis and a relative time scale in years on x axis (Fig. 4). It will take a few years for the seeds to germinate and become established seedlings and take much more years for the seedlings to become established saplings in the forest stands without major disturbances.
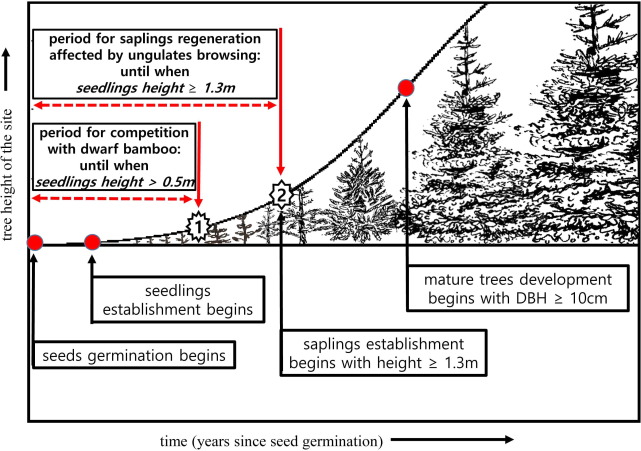
There are two important time periods for the successful regeneration of Korean fir trees on Mt. Hallasan. The first time period can be called as the ‘period for competition with dwarf bamboo’, which is the period for the seeds dispersed from the tree crown to the ground to be successful in getting into the soils, being able to establish the roots to soil layers, competing with other plant species including the dense foliage of dwarf bamboo for water, nutrients, light and space; and eventually being free from the competition with the dense dwarf bamboo shrubs, which will take more than 10 years at the forest stands. The second time period can be called as the ‘period for saplings regeneration affected by ungulate browsing’, which is even longer and vulnerable period for the saplings to deer browsing from the seed germinates to the established saplings with the height of breast (about 1.3–1.4 m above the ground). This time period can be more than 20 years at the forests.
Ungulate browsing: potentially the most powerful disturbing factor for sapling regeneration of the Korean fir
There are many threats awaiting for the germinating seeds to be developed into big seedlings and eventually into saplings for much longer time periods. Among many threats, ungulate browsing is quite fatal during any time from the germination of the seeds to the maturing seedlings and even to maturing saplings. In fact, browsing is the activity for the herbivores such as ungulate and livestock to get nutrients from the plant parts (Wilson, 1969). Here, ungulate is defined as “being hoofed” or “hoofed animal” including odd-toed ungulate such as horses and even-toed ungulate such as cattle, pigs, and deer. They are typically herbivorous (except for pigs which are omnivorous) and many of them are ruminants which employ specialized gut bacteria to allow them to digest cellulose which is not easily digested for many animals (Wikipedia, https://en.wikipedia.org/wiki/Ungulate).
In many places over the world, ungulate browsing in forests gives various effects on forest regeneration. Ungulate such as deer makes direct and negative effects on forest ecosystems causing the poor regeneration or complete interruption of conifers on the ecosystems of montane forests (Waller and Alverson, 1997; Rooney and Waller, 2003; Senn and Suter, 2003; Weisberg and Bugmann, 2003; Pépin et al., 2006; Takatsuki, 2009; Cutini et al., 2015; Swiss Federal Institute for Forest, 2015). Hidding et al. (2012) argued that the density of the ungulate is one of the determinants of the regeneration of the seedlings and saplings of balsam fir (Abies balsamea) in Canada. Côté et al. (2004) reviewed the ecological impacts of deer overabundance including the issues of plant tolerance and resistance to herbivory; plant community structure and interspecific competition; forest succession; ecosystem properties; cascading effects on animal species; dynamics and reversibility of deer impacts; and interactions with predators.
Siberian roe deer in Jeju Island has been loved by the people and was symbolized in the name of the crater lake, Baek-Rok-Dam (Korean, its meaning interpreted as White Deer Lake), at the top of Mt. Hallasan. Historically, Siberian roe deer population was almost wiped out from Hallasan by late 1970s–early 1980s by poaching and hunting by people. As the society changes, people became to realize the needs to restore the Siberian roe deer population on Jeju Island. Since the designation of the Mt. Hallasan as a National Park in Korea in 1970s, grazing of cattle and horses on Mt. Hallasan was prohibited in 1980s (Kang, 2013b). In contrast, as a measure for nature conservation in 1980s, Siberian roe deer was protected from nature including on Mt. Hallasan, Jeju Island. For the last three decades, the Siberian roe deer replaced the roles of cattle and horses grazing the plants on the sub-alpine forest ecosystem as well as many places on Jeju Island. As a result, the population size of the Siberian roe deer increased very rapidly without the existence of natural enemy such as wolf, which is an extinct animal from wild in Korean peninsula. On Mt. Hallasan, although there is no exact statistics on the density of Siberian roe deer, a study indicated that the population density is about 38 individuals/km2 in early 2000s on sub-alpine areas with elevation higher than 1600 m above mean sea level (Kim, 2002, 2006). Kim (2006) indicated that the increase in the number of Siberian roe deer appears to be responsible for turf destruction on Mt. Hallasan. In terms of the studies carried out on the population of Siberian roe deer in Jeju Island, there were studies on population dynamics (Kim and Hong, 2006; Jeon and Kim, 2011); peeling damage to tree saplings (Kim et al., 2012); home range and habitat studies by using wildlife tracking system (Kim et al., 2013); genetic diversity and population studies (Jo et al., 2012; Lee et al., 2015). It is interesting to note that Kim and Kim (2001) evaluated the native plant species on the preference of the leaves and needles by Siberian roe deer for their feeding. The results indicated that the Korean fir is one of the rarely fed species by Siberian roe deer. Despite this, extreme care should be taken not to extend to the argument that the seed germinates and small seedlings of Korean fir are also rarely fed by the Siberian roe deer on Mt. Hallasan, Jeju Island. Careful and similar studies are needed to test the preference or feeding tests from seed germinates to needles of young seedlings of different age groups by Siberian roe deer on the sites.
Here, the authors find a strong gap in knowledge on the interactions between ungulate animals and vegetation dynamics on Mt. Hallasan, Jeju Island, Korea. Further studies are needed on the herbivore-vegetation interactions in terms of the disturbance of the regeneration of Korean fir trees caused by the ungulate animals including the Siberian roe deer.
Dwarf bamboo: another powerful disturbing factor for sapling regeneration of the Korean fir
Another important consideration should be given to the potential for the dwarf bamboo to interrupt the germination of the seeds and to disturb the establishment of the seedlings at the stands, and, ultimately, to make big gaps in the frequency distribution of saplings observed at the forests on Mt. Hallasan, Jeju Island, Korea.
Around the world, it was well reported that dwarf bamboo produces inhibiting effects on the regeneration of tree species in the emergence, survival and growth of tree seedlings of firs as well as other tree species by the physical hindrance with dense foliage and root mates as well as the shade effects (Peters et al., 1992; Nakashizuka, 1988; Abe et al., 2001, 2002; Taylor et al., 2004, 2006; Doležal et al., 2009). In Korea, Song et al. (2014) indicated that the existence of dwarf bamboo is the major factor that limits the distribution of the seedlings of Korean fir on Mt. Hallasan.
To assure the success of the regeneration of the Korean fir trees outcompeting dwarf bamboo with dense foliage and root system, there are series of stages to check at the sites as below:
- –
- Stage I: Fertile cones and seeds ripening;
- –
- Stage II: High vitality of seeds after the dispersal from the cones;
- –
- Stage III: Successful germination of the seeds from the ground;
- –
- Stage IV: Successful rooting of the small seedlings established to the soil layers and overcoming the competition with the dwarf bamboo population with dense root and foliage system occupying the layer over ground floor;
- –
- Stage V: Survival of the seedlings from many potential threats such as ungulate browsing, droughts, damping-off pathogens, etc.;
- –
- Stage VI: Advancement of the seedlings to the saplings, the next stage of trees in size and shape, at the forest stands.
From these, the Stages from I to III are related to the genetic and physiological characteristics of the tree species. The Stage IV is a very critical one inhibiting the regeneration of the Korean fir seedlings at an earlier stage and disturbing the balance of the biodiversity of plant community by suppressing other small herbaceous plants as well as small shrubs in heights smaller than the canopy of the dwarf bamboo. It is expected for the germinating seeds to take for more than 10 years in outgrowing over the competing layers of the dwarf bamboo and in entering to next stage of taller seedlings. Although it was not well studied in the case of the Korean fir trees, many of the tree species of the genus Abies (fir trees) are shade tolerant tree species and have the strategy in growing slowly during earlier stages of their life span. So it can take more than 20 years for the germinating seeds to grow over the competition layer of the dwarf bamboo and become the taller seedlings. It is needed for us to know the mechanisms of the seedling development and the interactions of the developing seedlings with the dwarf bamboo in the forest stands.
At the Stages IV and V, after the seedlings to survive from many potential threats and to advance to the saplings, the saplings undergo severe competition with other deciduous hardwood species including Korean cherry, Silverly mountain ash, Erman’s Betula, Korean whitebeam and Mongolian oak, not to mention with another important coniferous tree species, i.e., Japanese yew. This indicates that the Korean fir trees should overcome additionally severe competition with other deciduous hardwood species before being developed into the mature trees of Korean fir at the forest stands.
Measures to take and further studies to be carried out
Studies indicated that mores studies are needed on the experiments with controlled grazing with exclosure experiments as well as on plant-herbivore interactions at a range of plant and animal densities (Hester et al., 2000; Kay and Bartos, 2000; Bottero et al., 2015). An example of the study on the interactions between deer and forest community is shown from an LTER (Long-Term Ecological Research) study site in Japan located at the Tomakomai Experimental Forest, Hokkaido University, Hokkaido, Japan (http://forestcsv.ees.hokudai.ac.jp/en/research/). Since 2004, the study team established a 20 ha study sites by installing fences as an experimental barrier for Sika deer to investigate the impacts of Sika deer on the community of plant and animal species. This experiment includes deer enclosure (16.5 ha), deer exclosure (2 ha) and control (1.5 ha) sites from an oak dominated forest. They have been studying the interactions between deer density and forest community including the effects on vegetation, birds, soil animals and arthropods. In addition, they incorporated the manipulation of the productivity of forest floor vegetation by the treatment of logging canopy trees and fertilization of the stands.
In addition, further mechanisms to promote the development of seedlings by applying various treatments including the removal of foliage and roots of dwarf bamboo with mechanical removal measures from the forest stands for consecutive years and/or the application of controlled burning or prescribed burnings and/or other measures could be designed and tested in terms of experiments.
Before having the reliable information on the disturbed regeneration of the seedlings and saplings of the Korean fir forests, efforts could be made to promote the regeneration of them, which may include the experiments for disturbing the site to expose the soil promoting the regeneration of seedling recruitment (Narukawa and Yamamoto, 2002; Taylor et al., 2004; Doležal et al., 2009). In the meanwhile, the measures in establishing fences to exclude the ungulate browsing and to protect the germinating sprouts as well as seedlings of various sizes should be taken. In addition, full scale research on the promotion of the regeneration of the seedlings and saplings of the Korean fir forests should be carried out, which include: seed development from the cones and seed germination (Song et al., 2010); competition of the seedlings and saplings with other plant species under different environmental conditions (Taylor and Halpern, 1991; Takahashi et al., 2001; Dobrowolska, 2008; Pellerin et al., 2010); the interaction of deer in browsing and the existence of the dwarf bamboo (Itô and Hino, 2004); topographic factors including aspects and altitudinal gradient (Hong et al., 2008; Dang et al., 2013); climate changes (Kim and Kim, 2000; Cunningham et al., 2006; Kim, 2010; Kang, 2013a; Park et al., 2015).
It is quite important for the managers of the forests to make sure of the successful regeneration of the Korean fir tree species by paying careful attention to the various processes of the regeneration and the many factors disturbing the natural regeneration of the tree species including ungulate browsing and severe hindrance of the dwarf bamboo in preventing various stages of the regeneration processes for the long period of time.
Conclusions
From this study, the authors maintain that, currently, the regeneration of the saplings at the stands of Korean fir (an endemic plant species) has been disturbed severely on Mt. Hallasan for the last two or three decades, and most possible factors affecting the regeneration for the managers of the forests to consider were the ungulate including Siberian roe deer to browse the seed sprouts and seedlings and the rapidly spreading dwarf bamboo hindering the establishment of regeneration of the seedlings from the very initial stage of the stand establishments. It is important to notice that the magnitude of the disturbance for the regeneration of the saplings of Korean fir trees has been big and continued. If the disturbance of the regeneration of seedlings and saplings by ungulate animals and dwarf bamboo continues at the forest stands, the decline of the Korean fir forests is expected to be real in the near future. Promoting the regeneration of the seedlings and saplings is the most important activity in preventing the decline of the Korean fir forest and, consequently, in assuring the sustainability of the forest ecosystem on Mt. Hallasan, Jeju Island, Korea. Therefore, major measures to promote the natural regeneration of the tree species should be taken to find out the causal factors and the mechanisms to disturb the natural regeneration of the tree species; to take steps in managing the potential threats including the interruption of the regeneration by Siberian roe deer and the competing dwarf bamboo; to carry out major activities in monitoring the dynamics of the forest stands including the structure, function, and development of the ecosystem, more specifically, in the dynamics of population and community and landscape patterns. Further the dynamics of the biodiversity among the species of the forest and adaptation measures of the forest under the changing regime of climate should be investigated among others.
Acknowledgments
This study was supported by the Ministry of Environment (MOE), Republic of Korea, through the Korea Institute of Environmental Science as the “The Converging Technology Program (2013000210001, Development of a Sensor-Network Based Platform System Technology for Monitoring Sub-Alpine Ecosystems on Climate Change in Korea)” and also supported by research program of Kookmin University, Seoul, Republic of Korea.
References
- Abe et al., 2001 M. Abe, H. Miguchi, T. Nakashizuka An interactive effect of simultaneous death of dwarf bamboo, canopy gap, and predatory rodents on beech regeneration. Oecologia, 127 (2001), pp. 281-286
- Abe et al., 2002 M. Abe, J. Izaki, H. Miguchi, T. Masaki, A. Makita, T. Nakashizuka The effects of Sasa and canopy gap formation on tree regeneration in an old beech forest. J. Veg. Sci., 13 (2002), pp. 565-574
- Bottero et al., 2015 A. Bottero, F. Meloni, R. Motta Ungulate browsing exclusion: effects and medium-term trends in coppice woods in north-western Italy. Effects of ungulate browsing on forest regeneration and silviculture, Proceeding of IUFRO International Conference, 14–16 October 2015, Birmensdorf, Switzerland (2015)
- Côté et al., 2004 S.D. Côté, T.P. Rooney, J.P. Tremblay, C. Dussault, D.M. Waller Ecological impacts of deer overabundance. Annu. Rev. Ecol. Evol. Syst., 35 (2004), pp. 113-147
- Cunningham et al., 2006 C. Cunningham, N.E. Zimmermann, V. Stoeckli, H. Bugmann Growth of Norway spruce (Picea abies L.) saplings in subalpine forests in Switzerland: does spring climate matter?. For. Ecol. Manage., 228 (2006), pp. 19-32
- Cutini et al., 2015 A. Cutini, F. Chianucci, M. Apollonio Early and long-term impacts of browsing by roe deer in coppiced woods in Central Italy. Effects of ungulate browsing on forest regeneration and silviculture, Proceeding of IUFRO International Conference, 14–16 October 2015, Birmensdorf, Switzerland (2015)
- Dang et al., 2013 H. Dang, K. Zhang, Y. Zhang, X. Tong, Q. Zhang Regeneration dynamics of subalpine fir (Abies fargesii) forest across the altitudinal range in the Shennongjia Mountains, central China. J. Plant Ecol., 6 (1) (2013), pp. 36-47
- Dobrowolska, 2008 D. Dobrowolska Growth and development of silver fir (Abies alba Mill.) regeneration and restoration of the species in the Karkonosze Mountains. J. For. Sci., 54 (9) (2008), pp. 398-408
- Doležal et al., 2009 J. Doležal, S. Matsuki, T. Hara Effects of dwarf bamboo understory on tree seedling emergence and survival in a mixed-oak forest in northern Japan: a multi-site experimental study. Commun. Ecol., 10 (2) (2009), pp. 225-235
- Ducey, 2006 M.J. Ducey The reverse-J and beyond: developing practical, effective marking guides. Proceedings of Implementing Uneven-Aged Management in New England: Is It Practical. University of New Hampshire Cooperative Extension, Caroline A. Fox Research and Demonstration Forest, Hillsborough, NH. April 13, 2006 (2006)
- Halpern, 2001 C.B. Halpern Constraints on Conifer Regeneration: A Review of the Literature and a Conceptual Model for the Pacific Northwest region. HJ Andrews Experimental Forest Long Term Ecological Research (2006). http://andrewsforest.oregonstate.edu/pubs/webdocs/reports/regen.cfm?topnav=66
- Helms, 1998 J.A. Helms. The Dictionary of Forestry, Society of American Foresters (1998). 210p
- Hester et al., 2000 A.J. Hester, L. Edenius, R.M. Buttenschøn, A.T. Kuiters Interactions between forests and herbivores: the role of controlled grazing experiments. Forestry, 73 (4) (2000), pp. 381-391
- Hidding et al., 2012 B. Hidding, J.P. Tremblay, S.D. Côté Survival and growth of balsam fir seedlings and saplings under multiple controlled ungulate densities. For. Ecol. Manage., 276 (2012), pp. 96-103
- Hong et al., 2008 S.G. Hong, J.J. Kim, H.K. Cho Studies on natural regeneration of Abies koreana. Natl. Acad. Sci., 47 (1) (2008), pp. 71-84. (in Korean with English abstract)
- Itô and Hino, 2004 H. Itô, T. Hino Effects of deer, mice and dwarf bamboo on the emergence, survival and growth of Abies homolepis (Piceaceae) seedlings. Ecol. Res., 19 (2004), pp. 217-223
- Jeon and Kim, 2011 D.U. Jeon, D.H. Kim Application of an augmented predator-prey model to the population dynamics of roe deer in Jeju. Korean Syst. Dyn. Res., 12 (2) (2011), pp. 95-126
- Jo et al., 2012 Y.S. Jo, T.W. Kim, B.J. Choi, H.S. Oh Current status of terrestrial mammals on Jeju Island. J. Species Res., 1 (2) (2012), pp. 249-256
- Kang, 2013a H. Kang Ecological responses of plants to climate change: research trends and its applicability in Korea. Korean J. Environ. Ecol., 46 (3) (2013), pp. 319-331. (in Korean with English abstract)
- Kang, 2013b M. Kang Stock-farming folklife and extinction of Mt. Halla sangsan-graze during the modern and contemporary. Cheju Natl. Univ. Tamla Cult. Res., 43 (2013), pp. 137-195. (in Korean with English abstract)
- Kay and Bartos, 2000 C.E. Kay, D.L. Bartos Ungulate herbivory on Utah aspen: assessment of long-term exclosures. J. Range Manage., 53 (2000), pp. 145-153
- Kerr and Mackintosh, 2012 G. Kerr, H. Mackintosh Long-Term survival of saplings during the transformation to continuous cover. Forests, 3 (2012), pp. 787-798
- Kerr et al., 2002 G. Kerr, B. Mason, R. Boswell, A. Pommerening Monitoring the transformation to continuous cover management. Forestry Commission Information Note 45, Forestry Commission, Edinburgh, UK (2002)
- Kim, 1994 E.S. Kim Decline of tree growth and the changes of environmental factors on high altitude mountains. A Report to Korea Science and Engineering Foundation (KOSEF 921–1500-018-2) (1994)
- Kim, 1996 Y.S. Kim Study on the Change of Forest Type and the Diameter Growth Characteristics of Korean Fir (Abies koreana Wilson) on Mt. Halla. (A Thesis for Master of Science). Graduate School, Kookmin University, Seoul, Korea (1996). p. 42
- Kim, 2002 M.H. Kim Turf exfoliation in the subalpine zone of Mt. Halla. J. Geomorphol. Assoc. Korea, 9 (1) (2002), pp. 71-81. (in Korean with English abstract)
- Kim, 2006 M.H. Kim Rates and processes of bare patch denudation in the subalpine grassland of Mt. Halla. J. Korean Geog. Soc., 41 (6) (2006), pp. 657-669. (in Korean with English abstract)
- Kim, 2010 E.S. Kim Statistical interpretation of climate change in Seoul, Korea, over the last 98 years. J. Ecol. Field Biol., 33 (1) (2010), pp. 37-45
- Kim and Choo, 2000 G.T. Kim, G.C. Choo Comparison of growth condition of Abies koreana Wilson by districts. Korean J. Env. Ecol., 14 (1) (2000), pp. 80-87. (in Korean with English abstract)
- Kim and Hong, 2006 D.H. Kim, Y.K. Hong A systemic approach for roe deer in Jejudo. Korean Syst. Dyn. Res., 7 (2) (2006), pp. 191-213
- Kim and Kim, 2000 E.S. Kim, Y.S. Kim Possibility of climate change and simulation of soil moisture content on Mt. Hallasan National Park, Chejudo Island, Korea. Korean J. Ecol., 23 (2) (2000), pp. 117-123
- Kim and Kim, 2001 J.E. Kim, M.H. Kim Study on the native plants preferably fed by the roe deer (Capreolus pygargus tianschanicus) in Mt. Halla. Cheju Natl. Univ. J. Basic Sci., 14 (1) (2001), pp. 63-72. (in Korean with English abstract)
- Kim et al., 2007a B.S. Kim, J.G. Oh, H.S. Oh The current number of its individuals and management plan of roe deer (Capreolus pygargus tianschanicus Satunin) inhabiting Mt. Halla (Hallasan), Jeju Island, Korea. Korean J. Env. Ecol., 21 (4) (2007), pp. 366-373. (in Korean with English abstract)
- Kim et al., 2007b G.T. Kim, G.C. Choo, T.W. Um Studies on the structure of Abies koreana community at subalpine zone in Hallasan. Korean J. Env. Ecol., 21 (2) (2007), pp. 161-167. (in Korean with English abstract)
- Kim et al., 2012 E.M. Kim, Y. Park, J.O. Kwon, J.E. Kim, C.W. Kang, C.B. Lee Peeling damage of sapling caused by the developing process of roe deer antlers in warm-temperate forests of Jeju Island. Korean J. Agric. For. Meteorol., 14 (4) (2012), pp. 254-259. (in Korean with English abstract)
- Kim et al., 2013 E.M. Kim, J.O. Kwon, C.W. Kang, K.M. Song, D.W. Min Home range size and habitat environment related to the parturition of roe deer at warm-temperate forest in Jeju Island using GPS-CDMA based wildlife tracking system. J. Korean Assoc. Geog. Inf. Stud., 16 (2) (2013), pp. 65-74. (in Korean with English abstract)
- Koh et al., 1996 J.G. Koh, D.S. Kim, S.C. Koh, M.H. Kim Dynamics of Abies koreana forests. Cheju-Do Isl. Res., 13 (1996), pp. 223-241. (in Korean with English abstract)
- Kohyama, 1988 T. Kohyama Etiology of “Shimagare” dieback and regeneration in subalpine Abies forests of Japan. GeoJournal, 17 (2) (1988), pp. 201-208
- Korea Forest Research Institute, 2015 Korea Forest Research Institute Conservation of Korean fir forests on Mt. Hallasan, Jeju Island, Korea. Proceedings of the International Symposium on the Conservation of Korean fir forests on Mt. Hallasan, Jeju Island, Korea, Warm Temperate and Subtropical Forest Research Center of the Korea Forest Research Institute, Seogwipo City, Jeju Island, Korea (2015)
- Leak, 1996 W.B. Leak Long-term structural change in uneven-aged northern hardwoods. For. Sci., 42 (2) (1996), pp. 160-165
- Lee et al., 2010 B.Y. Lee, G.H. Nam, J.H. Yun, G.Y. Cho, J.S. Lee, J.H. Kim, K.H. Oh Biological indicators to monitor responses against climate change in Korea. Korean J. Plant Taxon., 40 (2010), pp. 202-207
- Lee et al., 2015 Y.S. Lee, N. Markov, I. Voloshina, A. Argunov, D. Bayarlkhagva, J.G. Oh, Y.S. Park, M.S. Min, H. Lee, K.S. Kim Genetic diversity and genetic structure of the Siberian roe deer (Capreolus pygargus) populations from Asia. BMC Genet., 16 (1) (2015), pp. 1-15
- Loewenstein et al., 2000 E.F. Loewenstein, P.S. Johnson, H.E. Garrett Age and diameter structure of a managed uneven-aged oak forest. Can. J. For. Res., 30 (2000), pp. 1060-1070
- Lorimer, 1980 C.G. Lorimer Age structure and disturbance history of a southern Appalachian virgin forest. Ecology, 61 (5) (1980), pp. 1169-1184
- Meyer, 1952 H.A. Meyer Structure, growth, and drain in balanced uneven-aged forests. J. For., 50 (2) (1952), pp. 85-92. Washington
- Nakashizuka, 1988 T. Nakashizuka Regeneration of beech (Fagus crenata) after the simultaneous death of undergrowing dwarf bamboo (Sasa kurilensis). Ecol. Res., 3 (1988), pp. 21-35
- Narukawa and Yamamoto, 2002 Y. Narukawa, S. Yamamoto Effects of dwarf bamboo (Sasa sp.) and forest floor microsites on conifer seedling recruitment in a subalpine forest, Japan. For. Ecol. Manage., 163 (2002), pp. 61-70
- Park et al., 2015 H.C. Park, J.H. Lee, G.G. Lee, G.J. Um Environmental features of the distribution areas and climate sensitivity assessment of Korean fir and Khinghan Fir. J. Environ. Impact Assess., 24 (3) (2015), pp. 260-277. (in Korean with English abstract)
- Pellerin et al., 2010 M. Pellerin, S. Saïd, E. Richard, J.L. Hamann, C. Dubois-Coli, P. Hum Impact of deer on temperate forest vegetation and woody debris as protection of forest regeneration against browsing. For. Ecol. Manage., 260 (2010), pp. 429-437
- Pépin et al., 2006 D. Pépin, P.C. Renaud, Y. Boscardin, M. Goulard, C. Mallet, F. Anglard, P. Ballon Relative impact of browsing by red deer on mixed coniferous and broad-leaved seedlings – an enclosure-based experiment. For. Ecol. Manage., 222 (2006), pp. 302-313
- Peters et al., 1992 R. Peters, T. Nakashizuka, T. Ohkubo Regeneration and development in beech-dwarf bamboo forest in Japan. For. Ecol. Manage., 55 (1992), pp. 35-50
- Rooney and Waller, 2003 T.P. Rooney, D.M. Waller Direct and indirect effects of white-tailed deer in forest ecosystems. For. Ecol. Manage., 181 (2003), pp. 165-176
- Rubin et al., 2006 B.D. Rubin, P.D. Manion, D. Faber-Langendoen Diameter distributions and structural sustainability in forests. For. Ecol. Manage., 222 (2006), pp. 427-438
- Senn and Suter, 2003 J. Senn, W. Suter Ungulate browsing on silver fir (Abies alba) in the Swiss Alps: beliefs in search of supporting data. For. Ecol. Manage., 181 (2003), pp. 151-164
- Song et al., 2010 J.H. Song, K.H. Jang, S.D. Hur Variation of seed and germination characteristics of natural populations of Abies koreana Wilson, a Korean endemic species. J. Korean For. Soc., 99 (6) (2010), pp. 849-854. (in Korean with English abstract)
- Song et al., 2014 K.M. Song, Y.J. Kang, H.J. Hyeon Vegetation structure at the slope direction and characteristic of seedlings of Abies koreana in Hallasan Mountain. J. Environ. Sci. Int., 23 (1) (2014), pp. 39-46. (in Korean with English abstract)
- Sprugel, 1976 D.G. Sprugel Dynamic structure of wave-regenerated Abies balsamea forests in the northeastern United States. J. Ecol., 64 (1976), pp. 889-911
- Swiss Federal Institute for Forest, 2015 Swiss Federal Institute for Forest Effects of ungulate browsing on forest regeneration and silviculture. Proceeding of IUFRO International Conference, 14–16 October 2015, Birmensdorf, Switzerland (2015)
- Takahashi et al., 2001 K. Takahashi, K. Homma, V.P. Vetrova, S. Florenzev, T. Hara Stand structure and regeneration in a Kamchatka mixed boreal forest. J. Veg. Sci., 12 (2001), pp. 627-634
- Takatsuki, 2009 S. Takatsuki Effects of sika deer on vegetation in Japan: a review. Biol. Conserv., 142 (2009), pp. 1922-1929
- Taylor and Halpern, 1991 A.H. Taylor, C.B. Halpern The structure and dynamics of Abies magnifica forests in the southern Cascade Range, USA. J. Veg. Sci., 2 (1991), pp. 189-200
- Taylor et al., 2004 A.H. Taylor, J. Huang, S.Q. Zhou Canopy tree development and undergrowth bamboo dynamics in old-growth Abies–Betula forests in southwestern China: a 12-year study. For. Ecol. Manage., 200 (2004), pp. 347-360
- Taylor et al., 2006 A.H. Taylor, S.W. Jang, L.J. Zhao, C.P. Liang, C.J. Miao, J. Huang Regeneration patterns and tree species coexistence in old-growth Abies–Picea forests in southwestern China. For. Ecol. Manage., 223 (2006), pp. 303-317
- Waller and Alverson, 1997 D.M. Waller, W.S. Alverson The white-tailed deer: a keystone herbivore. Wildl. Soc. Bull., 25 (2) (1997), pp. 217-226
- Wang et al., 2000 J.R. Wang, T. Letchford, P. Comeau, K.P. Kimmins Above- and below-ground biomass and nutrient distribution of a paper birch and subalpine fir mixed-species stand in the Sub-boreal Spruce zone of British Columbia. For. Ecol. Manage., 130 (2000), pp. 17-26
- Weisberg and Bugmann, 2003 P.J. Weisberg, H. Bugmann Forest dynamics and ungulate herbivory: from leaf to landscape. For. Ecol. Manage., 181 (2003), pp. 1-12
- Westphal et al., 2006 C. Westphal, N. Tremer, G. von Oheimb, J. Hansen, K. von Gadow, W. Härdtle Is the reverse J-shaped diameter distribution universally applicable in European virgin beech forests?. For. Ecol. Manage., 223 (2006), pp. 75-83
- Wilson, 1920 E.H. Wilson Four new conifers from Korea. J. Arnold Arboretum, 1 (1920), pp. 186-190
- Wilson, 1969 A.D. Wilson A review of browse in the nutrition of grazing animals. J. Range Manage. Arch., 22 (1) (1969), pp. 23-28
- Woo et al., 2013 K.S. Woo, Y.K. Sohn, U.S. Ahn, S.H. Yoon, A. Spate Jeju Island Geopark – A Volcanic Wonder of Korea. Springer-Verlag, Berlin (2013)